materi reproduksi ternak
Hafez (2000) menyatakan bahwa kebuntingan adalah
suatu periode fisiologis pasca perkawinan ternak betina yang menghasilkan
konsepsi yang diikuti proses perkembangan embrio kemudian fetus hingga
terjadinya proses partus yang berlangsung sekitar 278 hari. Perkembangan
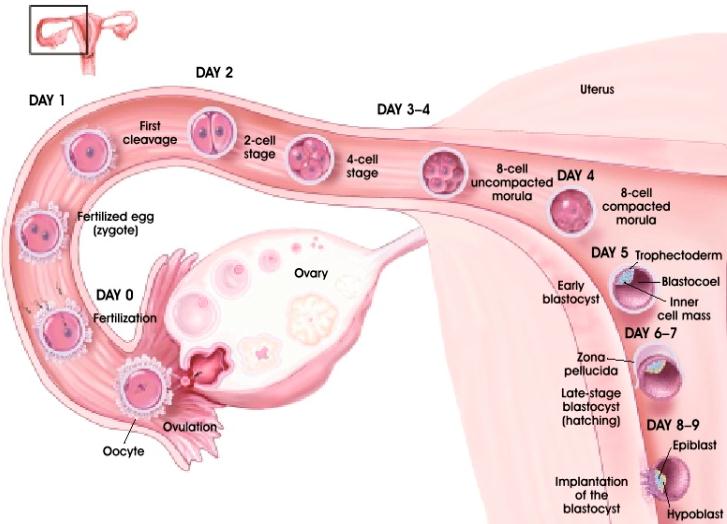
individu baru selama periode kebuntingan dibagi dalam (1) periode ovum yaitu
periode dari sejak terbentuknya zigote, morula dan blastula hingga implantasi
yang berlangsung antara 0-13 hari, (2) periode embrio yaitu periode dari
perkembangan blastula hingga pembentukan sistem organ termasuk plasenta yang
berlangsung antara 13-45 hari dan (3) periode fetus yaitu periode dari
pembentukan sistem organ dan plasenta hingga partus yang berlangsung dari 45
hari hingga partus (McDonald, 1975; Peters dan Ball 1987).
Keberhasilan kebuntingan sangat ditentukan oleh
beberapa proses penting di antaranya (1) folikel harus memiliki kemampuan
menghasilkan sel telur yang mampu dibuahi dan mengalami perkembangan embrionik,
(2) lingkungan oviduk dan uterus harus memiliki kelayakan untuk pengangkutan
gamet, fertilisasi dan perkembangan embrio dan (3) corpus luteum harus mampu
memelihara kebuntingan (Breuel, dkk., 1993). Sesaat setelah ovulasi maka sel
telur akan segera masuk ke tuba fallopii melalui infundibulum. Secara
berangsur-angsur perubahan fisiologi akan terjadi yaitu 8 jam setelah ovum
mengalami fertilisasi dan embrio akan menuju uterus untuk menyiapkan
perkembangan selanjutnya. Pembentukan membran plasenta sudah mulai terbentuk
pada 15-17 hari setelah fertilisasi yang merupakan periode Maternal Recognation
of Pregnancy dan bertujuan untuk mencegah pelepasan prostaglandin F2α dalam
melisiskan corpus luteum sehingga keberadaan progesteron dapat dipertahankan
dalam memelihara kebuntingan (Call, 1989; Beverly dan Sprott, 2004).
Beverly dan Sprott (2004) menyatakan bahwa
kebuntingan dapat ditentukan dalam tiga tahap. Tahap pertama meliputi tahap
kebuntingan 30-35 hari; 45 hari; 60 hari dan 90 hari. Kondisi embrio 30-35 hari
kebuntingan memiliki panjang sekitar ½ inchi dan terdapat gelembung seperti
balon yang berisi cairan dengan diameter ¾ inchi menyelimuti embrio. Usia
kebuntingan 45 hari, cornua uteri berisi fetus yang memiliki panjang sekitar 1
inchi. Membran luar dari dinding uterus berisi cairan dan adanya pertautan
antara karunkula dengan kotiledon dari membran fetus. Usia kebuntingan 60 hari,
cornua uteri yang dihuni oleh fetus nampak membesar hingga mencapai diameter
2½-3½ inchi dan panjang 8-10 inchi. Hal tersebut akan menarik uterus ke dalam
rongga tubuh hingga mencapai bagian pinggir dari pelvis.
Meningkatnya panjang fetus hingga mencapai 6½
inchi dan semakin beratnya beban uterus serta pembesaran pembuluh darah arteri
uterus adalah merupakan karakteristik usia kebuntingan sudah mencapai 90 hari.
Tahap kedua adalah kebuntingan 120 hari yaitu ukuran kepala sudah sebesar buah
lemon, diameter arteri uterus mencapai ¼ inchi dan kotiledon lebih nyata dengan
panjang sekitar 1½ inchi serta cornua uteri berdiameter 2-2½ inchi. Tahap
ketiga adalah usia kebuntingan sudah mencapai lebih dari 5 bulan dan usia
tersebut maka cornua uteri semakin masuk ke dalam rongga tubuh. Sejak usia
kebuntingan 6 bulan hingga melahirkan maka ukuran fetus, arteri uterus dan
kotiledon teraba lebih besar.
Untuk menyatakan keberhasilan kebuntingan dapat
dilakukan beberapa langkah di antaranya (1) nonreturn rate yaitu mengamati
tidak kembali estrus pada 18-24 hari pasca inseminasi, (2) palpasi rektal yaitu
melakukan perabaan dengan tangan melalui rektum terhadap perubahan organ
reproduksi pada 40-60 hari pasca inseminasi, (3) mengukur konsentrasi
progesteron melalui air susu atau plasma darah pada 21-24 hari pasca
inseminasi, (4) mammogenesis yaitu mengamati perkembangan kelenjar susu pada
usia kebuntingan 4 bulan (Peters dan Ball, 1987), (5) early conception factor
test yaitu mengetahui kebuntingan melalui glikoprotein dalam serum darah pada
48 jam pasca inseminasi dan (6) Ultrasonography yaitu alat yang digunakan dalam
mengetahui kebuntingan melalui gelombang suara pada 9 atau 12 hari pasca inseminasi
(Broaddus dan de Vries, 2005). Hasil penelitian menunjukkan bahwa teknik untuk
mengetahui kebuntingan dini yang memiliki tingkat akurasi tinggi adalah palpasi
rektal dan ultrasonography walaupun memerlukan waktu agak lama (Broaddus dan de
Vries, 2005).
pemeriksaan kebuntingan
Suatu pemeriksaan kebuntingan secara tepat dan
dini sangat penting bagi program pemulia-biakan ternak. Kesanggupan untuk
menentukan kebuntingan secara tepat dan dini perlu dimiliki oleh setiap dokter
hewan lapangan atau petugas pemeriksa kebuntingan. Palpasi perektal terhadap
uterus, ovarium dan pembuluh darah uterus adalah cara diagnosa kebuntingan yang
paling praktis dan akurat pada sapi. Pada pemeriksaan perektal tangan
dimasukkan kedalam inlet pelvis dan dengan telapak tangan yang membuka kebawah,
tangan digerakkan kesamping, keatas dan ke sisi lain. Apabila tidak ada
struktur yang teraba, uterus berada di lantai pelvis, cervik atau uterus teraba
di tepi pelvis pada sapi tua. Cervik yang keras dan ketat mudah dilokalisir pada
lantai pelvis atau di kranialnya. Korpus, kornu uteri dan ligamentum
interkornualis pada bifurcatio uteri dapat dipalpasi pada sapi yang tidak
bunting atau pada
kebuntingan muda. Ovarium dapat teraba dilateral
dan agak cranial dari cerviknya ( Toelihere, 1985).
Pengaturan hormonal pada kebuntingan.
Progesteron mempunyai peran dominan selama
kebuntingan terutama pada tahap-tahap awal. Apabila dalam uterus tidak terdapat
embrio pada hari ke 11 sampai 13 pada babi serta pada hari ke 15 – 17 pada
domba, maka PGF2α akan dikeluarkan dari endometrium dan disalurkan melalui pola
sirkulasi ke ovarium yang dapat menyebabkan regresinya corpus luteum (Bearden
and Fuquay, 2000). Apabila PGF2α diinjeksikan pada awal kebuntingan , maka
kebuntingan tersebut akan berakhir. Oleh sebab itu, embrio harus dapat
berkomunikasi tentang kehadirannya kepada sistem maternal sehingga dapat
mencegah PGF2α yang dapat menginduce luteolisis. Proses biokimia dimana embrio
memberi sinyal kehadirannya inilah yang disebut sebagai ” maternal recognition
of pregnancy”.
Pada sapi dan domba, unit embrionik memproduksi
suatu protein, yang disebut bovine interferon- λ dan ovine interferon- λ. Pada
kedua spesies tersebut, protein ini mempunyai perangkat antiluteolitik melalui
pengubahan biosintesa prostaglandin dan pengaturan reseptor uterin-oxytocin
(Gambar 1). Baik bovine interferon- λ pada sapi maupun ovine interferon- λ pada
domba, telah dilaporkan dapat menghambat sintesa
PGF2α dari endometrium. Pada domba, ovine
interferon- λ telah terbukti dapat meningkatkan konsentrasi PGE2 (sebuah hormon
antiluteolitik) dalam plasma darah pada kebuntingan hari ke 13. Sehubungan
dengan hal itu, apakah melalui peningkatan sintesa PGE2 atau penghambatan
sintesis PGF2α, rasio perbandingan yang tinggi antara PGE2 dan PGF2α adalah
kondisi yang mendukung pemeliharaan corpus luteum.
Konsentrasi tinggi progesteron, menurunkan tonus
myometrium dan menghambat kontraksi uterus. Efeknya pada myometrium tersebut,
membuat konsentrasi tinggi progesteron akan menghentikan siklus estrus dengan
mencegah dikeluarkannya gonadotropin. Progesteron diproduksi oleh corpus luteum
dan placenta.
Pada sapi, lutectomy ( pengambilan corpus luteum
atau injeksi PGF2α) pada kebuntingan tahap akhir, setelah 6 – 8 bulan
kebuntingan, tidak akan menyebabkan aborsi karena cukupnya steroid yang
diproduksi placenta. Pada domba, pengambilalihan fungsi placenta ini terjadi
pada 50 hari usia kebuntingan, sedang pada kuda sekitar 70 hari usia
kebuntingan. Pada beberapa spesies, ketika placenta mulai mengambil alih fungsi
sebagai sumber progesteron pada tahap dini kebuntingan, corpus luteum terus
mensekresi progesteron dan memelihara kebuntingan tersebut. Pregnancy-spesific
protein, protein B mungkin saja membantu corpus luteum kebuntingan pada sapi
dan domba (Bearden and Fuquay, 2000).
Polipeptida relaxin dan relaxin-like factors
yang diproduksi oleh corpus luteum (pada babi dan sapi) dan plasenta (pada
kuda) adalah penting selama terjadinya kebuntingan. Peran utamanya melunakkan
jaringan, yang menyebabkan otot-otot uterus dapat mengakomodir perkembangan
fetus. Relaxin menyebabkan saluran pelvis melebar, terutama pada tahap akhir
kebuntingan.
Konsentrasi estrogen rendah selama awal
kebuntingan dan meningkat pada pertengahan dan akhir kebuntingan. Pada kuda,
level estrogen cukup tinggi selama pertengahan kebuntingan. Sumber utama
estrogen ini adalah palsenta. Estrogen mengalami kenaikan yang progresif dalam
aliran darah uterus selagi proses kebuntingan terjadi. Estrogen bekerja sama
secara sinergis dengan progesteron pada perkembangan
dan persiapan kelenjar mammae untuk sintesa susu
setelah kelahiran. Laktogen plasenta juga sepertinya mempunyai peran dalam
perkembangan kelenjar mammae sebagaimana perannya dalam mengatur pertumbuhan
fetus.
3. Mengetahui proses partus, mekanisme partus
dan hormonal pada partus.
Fisiologi Kelahiran Sapi Perah Betina
Hafez (2000) menyatakan bahwa kelahiran atau
partus adalah proses pengeluaran fetus dan plasenta dari uterus induk sapi
bunting. Tahapan proses kelahiran meliputi (1) terjadi dilatasi cervix dan
masuknya fetus ke dalam cervix. Adanya pelunakan dan pelebaran cervix
menandakan bahwa fetus sudah berada di dalam cervix dan sekitar 2-6 jam lagi
maka induk akan partus, (2) pengeluaran fetus yang berlangsung sekitar 0,5-2 jam,
(3) pengeluaran plasenta yang umumnya berlangsung sekitar 3-5 jam pasca partus.
Kejadian pelunakan dan pelebaran cervix serta pengeluaran fetus pada sapi
paritas pertama memerlukan waktu yang lebih lama dibanding sapi yang sudah
beberapa kali partus (Bearden dan Fuquay, 1992).
McDonald (1975) dan Hafez (2000) menyatakan
bahwa mekanisme kelahiran merupakan peristiwa kompleks karena berhubungan
dengan proses endokrin, neural dan mekanik. Corpus luteum dari ovarium
merupakan sumber utama progesteron saat dimulainya kebuntingan karena
peranannya dalam memelihara kebuntingan. Namun saat akhir kebuntingan serta
seiring dengan membesarnya plasenta maka aktivitas corpus luteum semakin
menurun dan perannya digantikan oleh plasenta sebagai sumber progesteron (Shemesh,
2006).
Pada 20 hari hingga 2 hari menjelang partus,
konsentrasi progesteron secara berangsur mengalamai penurunan. Hal ini
disebabkan karena kortisol fetus merangsang plasenta menyekresikan estrogen
untuk menggertak peningkatan prostaglandin F2α dari endometrium uterus dan
merangsang peningkatan reseptor oxytocin dari endometrium. Kehadiran
prostaglandin F2α mampu menghambat sekresi progesteron dan merangsang
myometrium untuk berkontraksi. Kontraksi dari myometrium tersebut menghasilkan
refleks Ferguson sebagai akibat tekanan yang ditimbulkan oleh fetus terhadap
cervix dan vagina. Refleks Ferguson tersebut akan memberikan rangsangan
terhadap kelenjar hipofisa posterior untuk menyekresikan oxytocin. Oxytocin,
estrogen dan prostaglandin F2α bekerja secara simultan dalam memberikan
rangsangan kuat myometrium untuk berkontraksi sehingga terjadi proses pelepasan
perlekatan chorion fetus dari karunkula induk serta pengurangan volume uterus.
Pelepasan tersebut disebabkan adanya pengenduran perlekatan antara chorion
fetus dan karunkula karena adanya perobekan pembuluh darah sehingga darah lebih
banyak keluar (Peters dan Ball, 1987; Schmidt, 2005). Melalui mekanisme inilah
memungkinkan proses kelahiran berlangsung dari sejak terjadi dilatasi cervix
dan masuknya fetus ke dalam cervix, pengeluaran fetus hingga pengeluaran
plasenta.
4. Mengetahui proses post partus, laktasi dan
hormonnya.
Fisiologi Reproduksi Pasca Partus
1. Pengeluaran Plasenta
Plasenta merupakan penggabungan antara plasenta
foetalis (allantochorion) dan plasenta maternalis (endometrium) yang memiliki
fungsi fisiologis selama kebuntingan berlangsung. Pembentukan plasenta pada
awal kebuntingan merupakan membran fetus atau ekstra embrionik yang
berdiferensiasi ke dalam yolk sac, amnion, allantois dan chorion. Ternak sapi
memiliki tipe plasenta multiplek atau kotiledoner yaitu sebagian plasenta
maternal (karunkula) dan sebagian allantochorion (kotiledon) yang terletak
berhimpitan satu sama lain untuk membentuk plasentoma (Partodihardjo, 1980; dan
Hafez, 2000).
Hubungan antara kotiledon plasenta dengan
karunkula endometrium memiliki beberapa peran penting di antaranya (1)
melakukan pertukaran gas, (2) menyalurkan zat-zat makanan dari induk ke fetus,
(3) menyalurkan sisa-sisa metabolisme dari fetus ke sistem peredaran darah
induk dan (4) biosintesis sterol dengan Ca2+ sebagai second messenger dan
protein kinase C sehingga dihasilkan progesteron saat akhir kebuntingan melalui
fetal kotiledon (Bearden dan Fuquay, 1992; Stevenson, 2001; Shemesh, 2006).
Pengeluaran plasenta merupakan proses pemisahan
membran fetus dari karunkula induk dalam waktu beberapa jam pasca partus. Dalam
kondisi normal, membran fetus (plasenta) tersebut akan keluar dari tubuh induk
sekitar 6 – 12 jam pasca partus (Peters dan Ball, 1987; Hafez, 2000). Proses
pengeluaran plasenta dimulai dari terputusnya tali pusar yang menghubungkan
fetus dengan induk selama dalam kandungan sehingga mengakibatkan volume darah
dalam vili-vili turun dengan cepat. Semakin menurunnya volume darah dalam
pembuluh darah maka vili akan mengkerut dan volume uterus berangsur-angsur
menjadi kecil. Sekresi estrogen dan oxytocin yang merangsang kontraksi
myometrium akan menyebabkan terjadinya pengurangan volume uterus dan pelepasan
kripta-kripta endometrium tempat vili-vili plasenta bertaut dan secara bertahap
sisa plasenta dan tali pusar yang menggantung di mulut vulva akan menarik
plasenta secara keseluruhan keluar dari uterus (Partodihardjo, 1980).
Pengeluaran plasenta melebihi dari 12 jam maka
induk tersebut mengalami retensio secundinarum yaitu gangguan pengeluaran
plasenta (Hajurka, dkk., 2005). Hal tersebut dapat terjadi karena melemahnya
kontraksi myometrium sebagai akibat rendahnya kalsium dalam darah atau adanya
ketidakseimbangan hormon estrogen dan oxytocin serta partus sebelum waktunya
(Peters dan Ball, 1987; Goff dan Horst, 1997).
Paritas induk tidak memengaruhi performan
pengeluaran plasenta tetapi lebih banyak dipengaruhi oleh manajemen
pemeliharaan yang memengaruhi kondisi fisologis induk sapi. Kimura, dkk., (2002)
dan Epperson (2005) menyatakan bahwa sistem imunitas tubuh induk sangat
berpengaruh terhadap proses pengeluaran plasenta. Kandang tempat melahirkan
yang tidak memenuhi persyaratan kebersihan akan memberikan peluang kontaminasi
mikroorganisme terhadap organ reproduksi induk sehingga diperlukan perlawanan
dan sistem imunitas tubuh agar mikroorganisme yang mengganggu pengeluaran
plasenta dapat berkurang jumlahnya.
2. Pengeluaran Lochia
Lochia merupakan ekskretum cairan yang
mengandung runtuhan jaringan uterus yang bercampur dengan mukus, darah,
sisa-sisa membran fetus dan cairan fetus yang berwarna merah kecoklatan hingga
bening dan dikeluarkan pasca partus (Hafez, 2000 dan Amiridis, dkk., 2001,
Palmer, 2003). Hormon kortisol fetus merangsang plasenta menyekresikan estrogen
untuk menggertak peningkatan prostaglandin F2α dari endometrium uterus dan
merangsang peningkatan reseptor oxytocin dari endometrium. Kehadiran
prostaglandin F2α mampu menghambat sekresi progesteron dan merangsang
myometrium untuk berkontraksi. Kontraksi dari myometrium tersebut menghasilkan
refleks Ferguson sebagai akibat tekanan yang ditimbulkan oleh fetus terhadap
cervix dan vagina. Refleks Ferguson tersebut akan memberikan rangsangan
terhadap kelenjar hipofisa posterior untuk menyekresikan oxytocin. Oxytocin,
estrogen dan prostaglandin F2α bekerja secara simultan dalam memberikan
rangsangan kuat myometrium untuk berkontraksi sehingga menyebabkan runtuhnya
sel-sel endometrium dan bercampur dengan sekresi cairan uterus yang dihasilkan
oleh sel-sel kelenjar endometrium. Berlangsungnya proses kontraksi ritmik yang
diikuti pengeluaran runtuhan sel-sel endometrium dan sekresi cairan uterus
pasca partus menyebabkan pengeluaran lochia (Peters dan Ball, 1987; Schmidt,
2005).
Volume cairan yang dikeluarkan bergantung
paritas. Pluripara umumnya mengeluarkan lochia lebih banyak (1000-2000 ml)
dibandingkan primipara (kurang dari 500 ml). Pengeluaran lochia dalam volume
besar terjadi antara 1-3 hari dan berangsur menurun mulai hari ke-8. Sekitar
hari ke-9 biasanya keluar noda darah dan secara berangsur-angsur cairan akan
berwarna bening hingga berakhir antara 14-18 hari pasca partus. Meningkatnya
tekanan pembuluh darah arteri pada permukaan karunkula menyebabkan adanya
perembesan darah sehingga menimbulkan warna merah pada lochia. Lochia dalam
kondisi normal (tanpa kontaminasi bakteri) tidak akan mengeluarkan bau busuk
(Arthur, dkk., 1989).
Proses pengeluaran lochia bergantung pada
kecepatan pengeluaran plasenta dan keberadaan progesteron pasca partus (Arthur,
dkk., 1989 dan Thatcher, dkk., 2006). Induk pasca partus memiliki tiga tipe
fase luteal yaitu (a) pendek (4 -12 hari), (b) normal (13 – 20 hari) dan (c)
panjang (lebih dari 20 hari) (Terqui, dkk., 1982). Hal ini menunjukkan bahwa
keberadaan progesteron akan memengaruhi lamanya pengeluaran lochia karena
menurunnya estrogen akan mengurangi rangsangan kontraksi terhadap uterus. Suatu
penelitian membuktikan bahwa pada Pluripara umumnya mengeluarkan cairan lochia
lebih banyak (1000-2000 ml) dibandingkan primipara (kurang dari 500 ml) tetapi
primipara akan menyerap sebagian lochia ke dalam tubuh sehingga diperlukan
waktu bagi proses penyerapan tersebut. Suasana estrogenik dan frekuensi partus
yang lebih rendah pada primipara dibandingkan pluripara menyebabkan
terhambatnya proses penyerapan sebagian lochia sehingga pengeluaran lochia
primipara lebih lama (Arthur, dkk., 1989).
3. Estrus Pertama Pasca Partus
Secara umum diketahui bahwa berakhirnya estrus
yang diakhiri dengan proses ovulasi akan memberikan perubahan dalam konsentrasi
progesteron dalam darah. Konsentrasi progesteron sangat rendah (kurang dari
5ng/ml) terjadi pada 1-3 hari sebelum estrus kemudian akan mengalami
peningkatan secara drastis pada 4-12 hari pasca estrus dan pada 16-18 hari
konsentrasi progesteron akan konstan. Apabila ternak sapi tersebut bunting maka
keberadaan progesteron akan meningkat (sama atau lebih dari 5 ng/ml) guna
mempertahankan proses kebuntingan, tetapi progesteron akan segera menurun pada
18 hari siklus estrus hingga 2-4 hari sebelum estrus berikutnya berlangsung
(Grafik 1) (Rioux dan Rajotte, 2004).
Berlangsungnya proses kebuntingan akan
menyebabkan dipertahankannya corpus luteum untuk menghasilkan progesteron
hingga dua hari sebelum partus (Hendricks, 2004). Sejak 20 hari sebelum partus,
progesteron mulai berangsur menurun sebagai konsekuensi adanya rangsangan
kortisol (glukokortikoid) fetus untuk merangsang plasenta menyekresikan
estrogen guna menggertak prostaglandin F2α dari endometrium uterus serta
merangsang reseptor oxytocin dari endometrium. Kehadiran prostaglandin F2α
mampu menghambat sekresi progesteron dan merangsang myometrium untuk
berkontraksi (Peters dan Ball, 1987). Seiring penurunan progesteron, fetal
estrogen juga berperan dalam perejanan yang meliputi rangsangan myometrium
untuk berkontraksi serta relaksasi cervix dan vagina pada saat melahirkan,
pengeluaran fetus dan plasenta (McDonald, 1975; Goff dan Horst, 1997; Hafez,
2000; Shemesh, 2006). Di samping itu, estrogen akan merangsang peningkatan
hormon prolaktin yang berperan dalam laktogenesis. Keluarnya plasenta pasca
partus tidak menghilangkan keberadaan prostaglandin karena hormon tersebut akan
disekresikan oleh karunkula uterus induk hingga konsentrasinya menurun seiring
dengan menyusutnya ukuran uterus sekitar 14 hari pasca partus (Thatcher, dkk.,
2006).
Aktivitas ovarium selama kebuntingan
kecenderungan menurun terutama karena LH dalam darah yang rendah. Proses
folikulogenesis selama kebuntingan tidak dapat menghasilkan folikel dominan
karena tidak tersedianya LH untuk ruptura dinding folikel sehingga folikel
tersebut akan mengalami atresia. Kelenjar korteks adrenal yang menghasilkan
ACTH dan corticostreroid berperan dalam menekan sekresi LH. Peningkatan
pelepasan FSH dari hipofisa anterior pada 5-14 hari pasca partus menyebabkan
perkembangan folikel dalam ovarium (Tysseling, dkk., 1998 dan Rensis, 2001).
Setelah mengalami 2-3 kali atresia yang umumnya terjadi pada 7-8 hari
pencapaian folikel dominan selama siklus estrus berlangsung, maka folikel
tersebut baru dapat diovulasikan pada 18-24 hari (Grafik 2 dan 3) (Arthur,
dkk., 1989) dan Noseir, 2003) atau 21-24 hari pasca partus (Pate, 1999 dan
Rensis, 2001) tetapi umumnya ovulasi tersebut tidak disertai estrus karena
kurang memadainya ketersediaan estrogen dari induk (Sumber: Noseir, 2003).
Ukuran folikel dominan akan menentukan jumlah
sel granulosa dan produksi estrogen yang berperan dalam penampilan perilaku
estrus pertama pasca partus tetapi tidak dilakukan inseminasi karena proses
involusi uteri masih berlangsung (Fogwell, 1997). Gelombang folikel akan
menentukan lamanya siklus estrus pertama pasca partus. Hasil penelitian
menunjukkan bahwa folikulogenesis dengan dua gelombang memiliki potensi
konsentrasi estrogen yang lebih tinggi dan konsentrasi progesteron yang lebih
rendah dibandingkan folikulogenesis dengan tiga gelombang yang umum dialami
oleh induk sapi (Noseir, 2003). Kemampuan induk sapi primipara dan pluripara
dalam menampilkan folikel dominan yang akan diovulasikan dengan ukuran folikel
lebih dari 10 mm dan konsentrasi estrogen lebih dari 5,0 pg/ml sangat
bergantung pada tingkat keseimbangan energi pasca partus walaupun harus
kehilangan sebagian kondisi dan bobot tubuhnya.
Paritas induk tidak memengaruhi performan estrus
pertama pasca partus. Hasil penelitian menunjukkan bahwa induk sapi laktasi
(primipara dan pluripara) mempunyai kemampuan double ovulating (2,5±0,1)
dibandingkan sapi dara yang hanya single ovulating (1,0±0,0) sehingga estrogen
yang dihasilkan dapat melebihi dari 5,0 pg/ml (Wiltbank, dkk., 1995 dan Noseir,
2003). Kemampuan menghasilkan folikel dominan yang mampu diovulasikan lebih
dari satu (double ovulating) pada induk laktasi (primipara dan pluripara)
menyebabkan volume folikuler saat estrus meningkat sehingga terpenuhinya ketersediaan
estrogen bagi estrus dan kondisi tersebut teraktualisasi dalam intensitas
estrus. Sartori, dkk., (2002) mengungkapkan bahwa induk sapi laktasi (primipara
dan pluripara) mempunyai volume folikuler (2674,4±126,8 mm³) lebih besar
dibanding sapi dara (2202,8±168,5 mm³).
O’Connor dan Wu (2000) menggolongkan induk sapi
berdasarkan waktu ovulasi hingga terbentuknya corpus luteum pertama pasca
partus di antaranya: (1) kurang dari 40 hari, (2) 40-60 hari dan (3) lebih dari
60 hari. Semakin cepat terjadinya ovulasi pasca partus maka hipotalamus dan
hipofisa anterior akan menyekresikan gonadotropin guna kelangsungan
folikulogenesis. Secara hormonal, lamanya aktivitas luteal dapat diketahui dari
kadar progesteron yaitu bila kadar progesteron lebih dari 3 ng/ml (de Vries dan
Veerkamp, 2000).
4. Involusi Uteri
Involusi uteri adalah kembalinya ukuran dan
fungsi uterus dalam kondisi normal seperti sebelum mengalami kebuntingan
(Bearden dan Fuquay, 1992; Hafez, 2000). Kondisi tersebut dimulai sejak
berakhirnya minggu pertama pasca partus hingga involusi uteri terjadi secara
utuh yang ditandai oleh (1) menyusutnya ukuran corpus dan cornua uteri, (2)
uterus kembali berada di rongga pelvik, (3) konsistensi dan tekanan uterus
normal, (4) degenerasi karunkula yang diikuti oleh regenerasi jaringan epitel
uterus serta (5) terbebasnya cervix dari bakteri patogen (Morton, 2004 dan
Hajurka, dkk., 2005). Schirar dan Martinet (1982) menyatakan bahwa involusi
uteri umumnya terjadi melalui tiga proses yaitu (1) kontraksi, (2) pelepasan jaringan
dan (3) regenerasi jaringan.
Peningkatan prostaglandin F2α pada 7-23 hari
pasca partus akan memberikan rangsangan pada myometrium untuk melakukan
kontraksi (Kindahl, dkk., 1982). Proses pelepasan jaringan yang berlangsung
sekitar 15 hari pasca partus akan diikuti oleh penyusutan beberapa pembuluh
darah, regresi kelenjar uterus, penyusutan jumlah dan volume sel uterus.
Regenerasi sel epitel uterus dimulai segera setelah partus. Ruang di antara
karunkula akan diisi oleh sel-sel epitel yang baru pada 8 hari pasca partus dan
proses regenerasi secara keseluruhan akan berlangsung selama 4-5 minggu pasca
partus (Schirar dan Martinet, 1982).
Hendricks (2004) menyatakan bahwa selama
involusi uteri terjadi dua proses perubahan yaitu makroskopis dan mikroskopis.
Perubahan makroskopis meliputi proses penyusutan ukuran uterus yang diikuti
oleh pelepasan jaringan dan regenerasi endometrium. Penyusutan ukuran ini
disebabkan karena menguatnya kontraksi uterus yang terjadi setiap 3-4 menit
selama 24 jam pasca partus. Pada 5-10 hari pasca partus ukuran uterus sudah
mengalami penyusutan hingga 50 persen dan relaksasi cervix serta berakhirnya
pengelupasan massa karunkula. Sedangkan perubahan mikroskopis meliputi
perkembangan sel-sel epitel pada karunkula hingga mencapai 25 hari pasca partus
dan tidak adanya kontaminasi bakteri yang akan mengganggu perkembangan sel
epitel pada uterus.
Induk sapi pasca partus dalam kondisi normal
akan memberikan performan involusi yang signifikan dengan induk sapi yang
mengalami retensio sekundinarum, endometritis dan sistik ovari. Hasil
penelitian Hajurka, dkk. (2005) mengungkapkan bahwa performan involusi uteri
primipara kondisi normal (23,0±5,3 hari) dan yang mengalami kondisi tidak
normal (retensio sekundinarum, endometritis atau sistik ovari) (33,7±7,4 hari).
Sedangkan pada pluripara kondisi normal (27,3±5,5 hari) dan kondisi tidak
normal (37,3±8,2 hari). Namun primipara dalam kondisi normal memberikan
kontribusi performan involusi uteri (23,0±5,3 hari) tidak signifikan dengan
pluripara (27,3±5,5 hari). Hal ini menunjukkan bahwa paritas induk tidak
memengaruhi performan involusi uteri.
Peters dan Ball, (1987); Quintela, dkk., 2003
dan Hendricks (2004) menyatakan bahwa involusi uteri dipengaruhi oleh musim,
retensio sekundinarum, gangguan metabolik, hormon dan penyakit. Musim akan
memengaruhi involusi uteri yaitu induk sapi yang partus pada musim semi
(spring) dan panas (summer) akan mengalami waktu involusi uteri lebih pendek
dibandingkan musin dingin (winter). Gangguan metabolik yang memengaruhi
keterlambatan involusi uteri apabila kondisi induk sapi pasca partus mengalami
penurunan kadar kalsium darah (hypocalcemia) hingga mencapai kurang dari 2,0
mmol/L (Hendricks, 2004). Rendahnya konsentrasi estrogen akan berimplikasi
terhadap involusi uteri sehingga menyebabkan berkurangnya kontraksi myometrium
dan terjadi retensio sekundinarum (Peters dan Ball, 1987). Penyakit yang dapat
memengaruhi involusi uteri umumnya penyakit yang berhubungan dengan uterus
seperti pyometra dan endometritis (Quintela, dkk., 2003).
Proses pengamatan involusi uteri dapat dilakukan
melalui palpasi rektal, pemotongan ternak dan ultrasonography rektal
(Hendricks, 2004). Palpasi rektal untuk mengamati proses involusi uteri
merupakan cara yang umum dilakukan, yaitu dengan memerhatikan perubahan
struktur dan ukuran yang terjadi pada organ yang diamati pada setiap tahap
waktu. Selama tahap pertama (1-8 hari pasca partus) dapat diraba vagina dan
dilakukan penekanan pada uterus untuk mengamati kondisi cervix biasanya berada pada
tepi anterior dasar tulang pelvis. Permukaan uterus akan terasa keras serta
berkerut dan dalam kondisi rileks karunkula akan teraba melalui dinding uterus.
Dalam tahap kedua (8-10 hari pasca partus), uterus secara keseluruhan dapat
diraba. Struktur karunkula yang seperti kacang dan permukaan uterus akan terasa
halus, lunak dan berfluktuasi terutama pada cornua. Pada tahap ketiga ( 10-18
hari pasca partus) corpus uteri akan terasa seperti plastik lunak, tetapi
fluktuasi dari karunkula kurang dapat dirasakan, cervix nampak lebih kokoh dan
ukuran cornua terus mengecil hingga mendekati cornua sebelum bunting. Tahap
keempat (18-25 hari pasca partus) yang merupakan tahap akhir akan terasa adanya
peningkatan tekanan uterus dan pengurangan ukuran cornua sebelum bunting hingga
mendekati cornua uteri tidak bunting (Hendricks, 2004).
2.3.5. Estrus Kedua Pasca Partus dan Inseminasi
Buatan
Kehadiran performan estrus kedua pasca partus
menunjukkan bahwa induk sapi perah sudah mengalami keseimbangan energi positif
karena titik nadir keseimbangan energi negatif berlangsung hingga 3 minggu
pasca partus. Kondisi leptin, insulin dan IGF-I sudah berangsur mengalami
peningkatan kearah normal. Leptin dalam kondisi keseimbangan energi positif
akan meningkatkan feed intake yang diikuti peningkatan insulin dan IGF-I.
Jorritsma, dkk., (2003) menyatakan bahwa insulin dan IGF-I sangat memengaruhi
tingkat proliferasi folikel, produksi progesteron, estrogen sel granulosa dan
androgen sel theca. Insulin dan insulin-like growth factor-I (IGF-I) mampu
mengikat protein bagi kepentingan pertumbuhan folikel dan maturasi folikel
dominan sehingga terjadi peningkatan estrogen yang akan menimbulkan estrus
kedua pasca partus dan LH bagi kepentingan ovulasi.
Kontribusi leptin, insulin dan IGF-I dalam
metabolisme energi induk paritas I lebih diutamakan pada pemenuhan kebutuhan
pertumbuhan untuk mencapai kematangan fisiknya. Sedangkan induk paritas II dan
III lebih mengutamakan hasil metabolisme energi digunakan untuk memenuhi
kebutuhan pertumbuhan dan maturasi folikel ataupun produksi susu. Hasil
penelitian Wathes, dkk., (2005) mengungkapkan bahwa primipara memberikan
performan estrus kedua pasca partus (78,5±2,42 hari) lebih lama dibandingkan
pada pluripara (72,9±1,12 hari).
Performan estrus kedua pasca partus juga
menggambarkan uterus sudah mengalami involusi artinya secara fisiologis induk
mampu menerima kebuntingan berikutnya. Oleh sebab itulah, estrus kedua pasca
partus umumnya digunakan sebagai langkah awal dalam melakukan inseminasi buatan
pertama pasca partus. Inseminasi buatan adalah suatu proses mendeposisikan
semen ke dalam uterus (intrauterin) atau cervix (intracervical) dengan
menggunakan alat artificial copulation. Inseminasi buatan memberikan kontribusi
penting dalam days open dan calving interval karena keberhasilan inseminasi
pertama pasca partus yang menghasilkan kebuntingan akan memengaruhi lamanya
performan reproduksi tersebut.
Waktu yang tepat untuk inseminasi merupakan
dasar bagi deposisi semen ke dalam organ reproduksi induk. Pelaksanaan
inseminasi yang baik dilakukan pada 12-18 jam yang dihitung dari sejak awal
berlangsungnya estrus. .Penentuan waktu tersebut didasarkan pada kemampuan
spermatozoa dapat hidup dengan baik pada saluran reproduksi betina selama 18-24
jam, waktu ovulasi sel telur dan daya hidup sel telur untuk dapat dibuahi 10-20
jam. Sedangkan deposisi semen saat inseminasi dapat dilakukan pada corpus
uteri, cornua uteri bagian kanan dan cornua uteri bagian kiri (O’Connor dan
Peters, 2003). Indikator tingkat keberhasilan pelaksanaan inseminasi pada
estrus kedua pasca partus dapat diamati melalui service per conception dan
conception rate (Wells dan Burton, 2002).
DAFTAR PUSTAKA
Anonim. 2009. http://www6.ufrgs.br/favet/imunovet/molecular_immunology/blastocyst.jpg diakses Yogyakarta, 28 Desember 2009.
Bearden H.J. and W.J. Fuguay.R 2004. Applied
Animal Reroduction 6 th Ed. Philadelphia : Lea
& Febiger.
Arthur, dkk. 1989. Veterinary
Reproduction and Obstetrics. 6th Ed. Baliere Tindall.
Jainudeen, M.R. and Hafez. E.S.E. 2000.
Pregnancy Diagnosis, dalam Hafez, E.S.E and
Hafez, B. 2000. Reproduction in Farm
Animals. 7ed.. Philadelphia : Lippincott Williams &
Wilkins.
McDonald, L.E. 1971. Veterinary
Endocrinology and Reproduction. Philadelphia : Lea & Febiger.
Partodiharjo, S. 1987. Ilmu Reproduksi
Hewan. Jakarta : Mutiara Sumber Daya.
Salisbury, G.W, and VanDemark, N.L., 1985, Fisiologi
Reproduksi dan Inseminasi Buatan pada Sapi. Diterjemahkan oleh Djanuar, R.,
Yogyakarta : Gadjah Mada University Press.
Kebuntingan adalah serangkaian proses
fisiologis yang dimulai dari
terjadinya fertilisasi dan diakhiri dengan
kelahiran (Jainudeen & Hafez 2000).
Lama kebuntingan pada domba bervariasi
bergantung pada bangsanya yaitu
berkisar antara 144 – 153 hari (Johnson
& Everitt 2000; Senger 1999) dengan
rata-rata 148 hari.
Penanda Kebuntingan
Saat yang paling kritis dalam siklus
reproduksi ternak ditentukan oleh
kemampuan induk untuk menerima sinyal yang
dikirimkan oleh konseptus untuk
menghalangi terjadinya luteolisis dan
mempertahankan kebuntingan yang disebut
dengan maternal recognition of pregnancy
atau MRP (Senger 1999;
Geisert & Malayer 2000) . Pada
ruminansia sinyal penanda kebuntingan yang
utama adalah interferon tau (Johnson &
Everitt 2000; Spencer & Bazer 2004).
Progesteron merupakan hormon penjaga
kebuntingan. Keberadaan
progesteron di dalam uterus akan
menstimulir dan menjaga fungsi uterus sehingga
dapat dipergunakan untuk tempat
perkembangan embrio dini, implantasi,
plasentasi serta keberhasilan perkembangan
fetus dan plasenta sampai akhir masa
kebuntingan (Spencer et al. 2004).
Pada ternak domba, sudah dapat dinyatakan
bunting jika konsentrasi
progesteron dalam darah minimal 2,5 ng/ml
(Boscos et al. 2003) sedangkan
peneliti lainnya menyatakan bahwa
konsentrasi tertinggi progesteron pada fase
luteal pada 2-4 ng/ml dibandingkan dengan
saat estrus pada 1,5-0,8 ng/ml
(Ranilla et al. 1994). Peningkatan yang
drastis dari 2-4 ng/ml menjadi 12-20
ng/ml terjadi pada kebuntingan hari ke
60-125 (Edqvist & Stabenfeldt 1980),
karena plasenta dan atau konseptus sudah
memproduksi progesteron. Hal yang 10
sama terlihat level progesteron meningkat
dengan konsentrasi tertinggi 16 ng/ml
(Johnson & Everitt 2000).
Korpus luteum domba memproduksi progesteron
dalam jumlah yang
relatif rendah pada 50 hari pertama
kebuntingan, tetapi setelah melewati masa ini
plasenta merespon terhadap lueteinizing
hormon maupun prolaktin, untuk
mempersiapkan diri sebagai sumber utama
progesteron sampai kebuntingan
berakhir. Hal ini menunjukkan bahwa
konsentrasi progesteron baru terukur
setelah hari ke-60 (Schoenecker et al.
2004).
Apabila plasenta telah berfungsi dengan
sempurna maka meskipun
dilakukan ovariektomi produksi hormon yang
menjaga kebuntingan tetap
disekresikan karena fungsinya telah
digantikan oleh plasenta (Gambar 3). Domba
apabila dilakukan ovariektomi setelah hari
ke-50 tidak akan menyebabkan
terjadinya abortus (Senger 1999; Johnson
& Everitt 2000).
Estrogen
Aktivitas utama estrogen adalah menunjukkan
tanda berahi saat estrus,
meningkatkan ukuran uterus, aliran darah
uterus, meningkatkan ekspresi reseptor
progesteron terhadap oksitosin, mendorong
perkembangan organ fetus,
menstimulir produksi protein hepar induk
serta meningkatkan massa jaringan
mammae dan adipose (Hirako et al. 2003;
Senger 1999; Johnson & Everitt 2000).
Estrogen merupakan hormon yang selain
diproduksi oleh ovarium juga
diproduksi oleh kotiledon fetus
bersama-sama dengan karunkula induk
(Teng et al. 2002). Salah satu produk
deteksi kebuntingan (DEEA Gestdect®)
dengan memanfaatkan ikatan fenol yang
terikat pada gugus estrogen dalam urin,
mempunyai mempunyai akurasi pada domba dan
sapi berturut-turut 60-70 % dan
90 % (Samsudewa et al. 2005). Hal ini
menunjukan bahwa estrogen terukur
dalam urin domba bunting maupun tidak
bunting
Kebuntingan adalah proses bersatunya sel
sperma dan sel telur yang akan
membentuk zigot dan kemudian menjadi embrio
dan fetus. Lama kebuntingan ternak
babi berkisar antara 111-117 hari atau
rata-rata 114 hari. Meskipun perkembangan
sejak pembuahan hingga kelahiran merupakan
suatu proses berkesinambungan, kebuntingan dianggap terdiri dari tiga fase, yaitu
fase preimplantasi, embrio, dan
fetus (Sihombing, 2006)


{kind=link}
Komentar
Posting Komentar